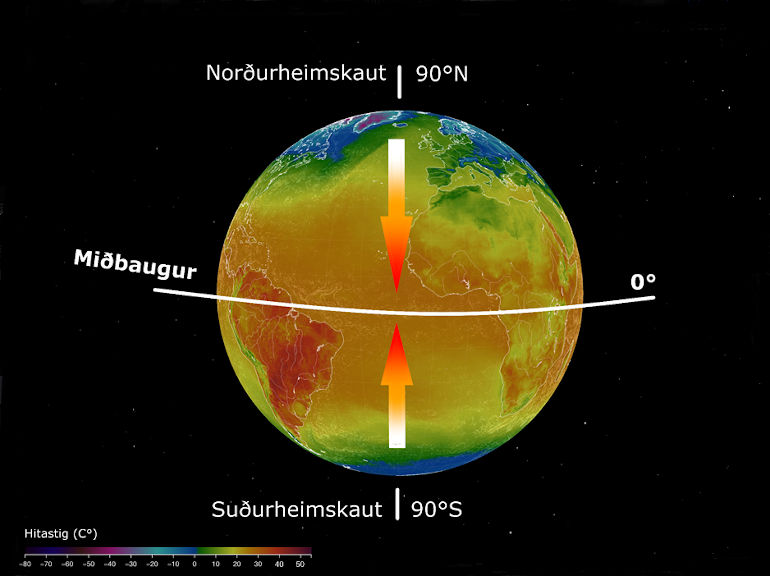
Mynd 1. Myndræn skýring á hugtakinu margbreytileikastigull miðbaugs. Örvar verða rauðari eftir því sem tegundafjöldi eykst.

Mynd 2. Köngulóin og liðfætlan Macracantha arcuata; þýtt lauslega sem ,,langhyrnubugða’’. Hún finnst í regnskógum Taílands og Malasíu og er eina tegund ættkvíslarinnar. Fyrst skráð sem Arana arcuata, síðar þekkt sem Gasteracantha arcuata.

Mynd 3. Fjórar tegundir faga af ættkvíslinni Kayflunavirus. Einangraðir úr regnskógi í Penang-þjóðgarðinum í Malasíu. Tegundirnar sýkja allar Salmonella-bakteríur, sem vekja vonir um að þær megi nýta til að vinna gegn smiti sem berst með vatni.
- Understanding Evolution. Settu þig í spor vísindamanna og skoðaðu margbreytileikastigul miðbaugsins með eigin augum!
- Rainforest Adventure. Heimsókn í Amasonregnskóg - gagnvirkur, gamaldags vafraleikur þar sem þú þarft að leysa gátur innan lífríkis regnskógarins.
- Agnarsson, I., Coddington, J.A. og Kuntner, M. (2013). Systematics – progress in the study of spider diversity and evolution. Í D. Penney (ritstjóri) Spider Research in the 21st Century (bls. 58-111). Siri Scientific Press.
- Aime, M.C., og Brearley, F.Q. (2012). Tropical fungal diversity: closing the gap between species estimates and species discovery. Biodiversity and Conservation, 21(9), 2177–2180. DOI: 10.1007/s10531-012-0338-7
- Basset, Y., Cizek, L., Cuénoud, P., Didham, R. K., Guilhaumon, F., Missa, O., Novotny, V., Ødegaard, F., Roslin, T., Schmidl, J., Tishechkin, A.K., Winchester, N.N., Roubik, D. W., Aberlenc, H.-P., Bail, J., Barrios, H., Bridle, J. R., Castaño-Meneses, G., Corbara, B., … Leponce, M. (2012). Arthropod Diversity in a Tropical Forest. Science, 338(6113), 1481–1484. DOI: 10.1126/science.1226727
- Black, T., og Jassal, R. (2016). Evapotranspiration. Í E. Johnson & Y. Martin (ritstjórar), A Biogeoscience Approach to Ecosystems (bls. 292-344). Cambridge University Press. DOI: 10.1017/CBO9781107110632
- Burgin C.J., Colella J.P., Kahn P.L., Upham N.S. (2018). How many species of mammals are there? Journal of Mammalogy. 99(1): 1–14. DOI: 10.1093/jmammal/gyx147
- Butler, R.A. (2020, 14. 08). RAINFOREST INFORMATION. Mongabay. https://rainforests.mongabay.com/
- Coddington, J.A., Ingi Agnarsson, Miller, J.A., Kuntner, M. og Hormiga, G. (2009). Undersampling bias: the null hypothesis for singleton species in tropical arthropod surveys. Journal of Animal Ecology, 78(3), 573-584. DOI: 10.1111/j.1365-2656.2009.01525.x
- Erwing, T.L. (1982). Tropical Forests: Their Richness in Coleoptera and Other Arthropod Species. The Coleopterists Bulletin, 36(1), 74-75.
- Johnson, H.J. (2023, 19. október). Rainforest. National Geographic. https://education.nationalgeographic.org/resource/rain-forest/
- Mutusamy, P., Banga Singh, K. K., Su Yin, L., Petersen, B., Sicheritz-Ponten, T., Clokie, M. R. J., Loke, S., Millard, A., Parimannan, S., og Rajandas, H. (2023). Phenotypic Characterization and Comparative Genomic Analysis of Novel Salmonella Bacteriophages Isolated from a Tropical Rainforest. International Journal of Molecular Sciences, 24(4). DOI: 10.3390/ijms24043678
- Newman, A. (2002). Tropical Rainforest: Our Most Valuable and Endangered Habitat With a Blueprint for Its Survival into the Thir Millenium (2.útgáfa). Checkmark.
- Nogueira, W.G., Gois, B.V.A., Pinheiro, K.D.C., Aragão, A O., Queiroz, A.L.C., da Silva, A. L., Folador, A. C., og Ramos, R. T. J. (2022). Viral Metagenomics Reveals Widely Diverse Viral Community of Freshwater Amazonian Lake. Frontiers in public health, 10. DOI: 10.3389/fpubh.2022.869886
- Paula, F.S., Rodrigues, J.L. M., Zhou, J., Wu, L., Mueller, R.C., Mirza, B.S., Bohannan, B.J. M., Nüsslein, K., Deng, Y., Tiedje, J. M., og Pellizari, V. H. (2014). Land use change alters functional gene diversity, composition and abundance in Amazon forest soil microbial communities. Molecular ecology, 23(12), 2988-2999. DOI: 10.1111/mec.12786
- Rodrigues, J.L., Pellizari, V.H., Mueller, R., Baek, K., Jesus Eda, C., Paula, F.S., Mirza, B., Hamaoui, G.S. Jr., Tsai, S.M., Feigl, B., Tiedje, J.M., Bohannan, B.J. og Nüsslein, K. (2012). Conversion of the Amazon rainforest to agriculture results in biotic homogenization of soil bacterial communities. PNAS 110(3), 988-93. DIOI: 10.1073/pnas.1220608110
- Santos-Júnior, C.D., Santos-Júnior CD, Kishi LT, Toyama D, Soares-Costa A, Oliveira TC, de Miranda FP, Henrique-Silva F. (2017). Metagenome Sequencing of Prokaryotic Microbiota Collected from Rivers in the Upper Amazon Basin. Genome Announc, 5(2):e01450-16. DOI: 10.1128/genomea.01450-16
- Schlesinger, W.H. og Jasechko, S. (2014) Transpiration in the global water cycle. Agricultural and Forest Meteorology, 189–190, 115-117. DOI: 10.1016/j.agrformet.2014.01.011
- Schloss, P.D., og Handelsman, J. (2004). Status of the Microbial Census. Microbiology and Molecular Biology Reviews, 68(4), 686–691. DOI: 10.1128/mmbr.68.4.686-691.2004
- Schneider, D., Engelhaupt, M., Allen, K., Kurniawan, S., Krashevska, V., Heinemann, M., Nacke, H., Wijayanti, M., Meryandini, A., Corre, M. D., Scheu, S., og Daniel, R. (2015). Impact of Lowland Rainforest Transformation on Diversity and Composition of Soil Prokaryotic Communities in Sumatra (Indonesia). Frontiers in Microbiology, 6. 1-12. DOI: 10.3389/fmicb.2015.01339
- Schulz, G., Schneider, D., Brinkmann, N., Edy, N., Daniel, R., Polle, A., Scheu, S., og Krashevska, V. (2019). Changes in Trophic Groups of Protists With Conversion of Rainforest Into Rubber and Oil Palm Plantations. Frontiers in Microbiology, 10. DOI: 10.3389/fmicb.2019.00240
- Tedersoo, L. og Nara, K. (2009). General latitudinal gradient of biodiversity is reversed in ectomycorrhizal fungi. New Phytologist, 185(2), 351–354. DOI: 10.1111/j.1469-8137.2009.03134.x
- Tortosa, P. (2022). Introducing Microbes in the Context of the Madagascar Biodiversity Hotspot. Í: S. M. Goodman (ritstjóri) The New Natural History of Madagascar (bls. 265-267). Princeton University Press.
- Willig, M.R. og Presley, S.J .(2018). Latitudinal Gradients of Biodiversity: Theory and Empirical Patterns. Í D.A. Dellasala og M.I. Goldstein, (ritstjórar) Encyclopedia of the Anthropocene, 3 (bls. 13-19). DOI: 10.1016/B978-0-12-809665-9.09809-8
- Þóra Ellen Þórhallsdóttir. (1989). Regnskógar hitabeltisins. Náttúrufræðingurinn, 59(1), 9-37. https://timarit.is/gegnir/991006730589706886
- Mynd 1. Education and Outreach Group at the Byrd Polar and Climate Research Center (BPCRC), The Ohio State University (OSU). Fluid Earth, kennslutól sem sýnir loftslag, sæ og sífrera í rauntíma myndrænt. Mynd aðlöguð með paint.
- Mynd 2. Rushen. (2012). Macracantha arcuata - Curved Spiny Spider. Eigandi myndar: Thai National Parks. Flickr. Birt undir CC BY-SA 2.0 DEED. Sótt 25. nóvember 2023.
- Mynd 3. Mutusamy, P. o.fl. (2023). Phenotypic Characterization and Comparative Genomic Analysis of Novel Salmonella Bacteriophages Isolated from a Tropical Rainforest. International Journal of Molecular Sciences, 24(4). DOI: 10.3390/ijms24043678
Þetta svar var unnið í tengslum við námskeiðið Örverufræði II (LÍF201G) við Líf- og umhverfisvísindadeild Háskóla Íslands.


